Inbred – jak teoria stała się praktyką
Blisko 100 lat temu powstała teoria wyjaśniająca zjawisko chowu wsobnego (inbredu), dając podwaliny pod matematyczne obliczanie współczynników inbredu dla pojedynczych zwierząt oraz całej populacji. Pomimo upływu lat temat ten cieszy się niesłabnącą popularnością i zajmuje hodowców oraz naukowców na całym świecie. Dobre zrozumienie mechanizmu genetycznego, jaki stoi za zjawiskiem inbredu, pozwala na lepsze radzenie sobie z nim w codziennej pracy w stadzie.
tekst: dr inż. Magdalena Graczyk-Bogdanowicz
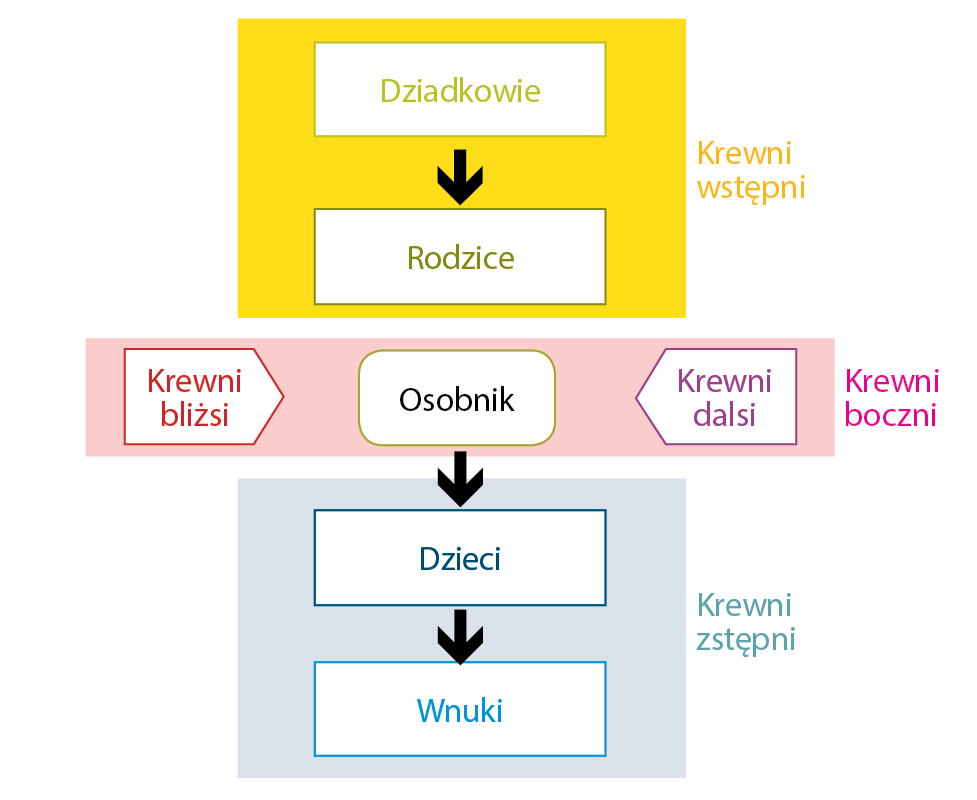
Żeby można było rozpocząć nasze rozważania na temat inbredu, należy wspomnieć o relacji, jaka może występować pomiędzy osobnikami, bo to właśnie z nią wiąże się inbred, oraz o źródłach, z jakich czerpiemy tę informację. Podstawowym źródłem informacji są rodowody w postaci ksiąg hodowlanych, na ich podstawie możemy określić zależność genetyczną pomiędzy osobnikami i rodzaj relacji. Spójrzmy na graf. Jakie możemy mieć typy krewnych?
Z krewnymi w linii prostej mamy bezpośrednią relację, są to wszyscy krewni wstępni i zstępni, tj. dziadkowie, rodzice, dzieci. Krewnymi w linii bocznej nazywamy wszystkie pozostałe osobniki, bliżej lub dalej spokrewnione. Relacja pomiędzy osobnikiem i krewnym bocznym ma charakter pośredni, tj. kuzynostwo, wujostwo i dalsi krewni. Pomiędzy wszystkimi zwierzętami przedstawionymi na grafie jesteśmy w stanie określić poziom relacji, a więc spokrewnienie. Definiowane jest ono jako podobieństwo genetyczne pomiędzy osobnikami, wynikające z posiadania takich samych genów, które odziedziczyły po jednym przodku bądź kilku wspólnych przodkach. To właśnie spokrewnienie jest podstawą rozważań o inbredzie.
Ale czym właściwie jest inbred z genetycznego punktu widzenia? W klasycznej teorii Fisher’a i Wright’a inbred jest przede wszystkim prawdopodobieństwem, że osobnik w konkretnym miejscu w genomie (locus) posiada dwa allele (fragmenty DNA), które są identyczne przez pochodzenie. To znaczy, że zostały odziedziczone od wspólnego przodka w wyniku kojarzenia krewniaczego jego rodziców. Dzieje się tak dlatego, że w trakcie segregacji materiału genetycznego do komórki jajowej lub plemnika (gamet) każdy rodzic przekazuje połowę genów. Losowo z każdej pary alleli do gamety trafia jedna kopia. W związku z powyższym potomek takiej pary odziedziczy dwa allele – jeden od matki, a drugi od ojca. Jeśli rodzice są ze sobą blisko spokrewnieni, istnieje duże prawdopodobieństwo, że oba allele pochodzą od tego samego osobnika – przodka obojga, a więc będą one identyczne, a miejsce w genomie staje się homozygotyczne. Homo z języka greckiego oznacza ‘taki sam’, dlatego też o inbredzie mówimy również jako o poziomie homozygotyczności, a więc frekwencji ‘jednakowych’ miejsc w całym DNA zwierzęcia. Należy wspomnieć, że nie każde miejsce w genomie, które jest homozygotyczne, zawiera allele, które są identyczne przez pochodzenie. Niektóre miejsca w DNA są jednakowe przez stan. To znaczy, że dwa allele są podobne strukturalnie, a miejsca takie są naturalnie rozmieszczone w całym materiale genetycznym. Zwykle są one rasowo lub gatunkowo specyficzne i występują u wszystkich zwierząt z danej populacji. Wracając do samego inbredu, objawia się on u potomka i wynika ze spokrewnienia pomiędzy jego rodzicami. Im wyższy poziom inbredu osobnika, tym bliższe było spokrewnienie pomiędzy rodzicami, ponieważ inbred jest definiowany jako połowa spokrewnienia jego rodziców. W populacji poddawanej selekcji wszystkie zwierzęta są ze sobą w mniejszym lub większym stopniu spokrewnione. Zgodnie z tym stwierdzeniem należałoby unikać kojarzenia osobników, które są ze sobą bardziej spokrewnione niż średnia dla całej populacji.
Dlaczego w takim razie mówimy w ogóle o prawdopodobieństwie, a nie o pewności w kontekście wyliczania klasycznego inbredu? Ponieważ informacją, na podstawie której obliczany jest współczynnik inbredu, jest rodowód, który pozwala z pewnym przybliżeniem określić, jakie geny mógł przekazać dany rodzic swojemu potomstwu. Jednak – co należy wspomnieć – klasyczne szacowanie inbredu na podstawie informacji rodowodowej jest wrażliwe na jakość i kompletność rodowodu. Jeśli informacja ta jest niekompletna lub niedokładna, wówczas inbred zwierzęcia może być niedoszacowany. Jeśli dysponujemy płytkim rodowodem, inbred danego osobnika może być bardzo niski lub nawet zerowy. Jednak zwiększenie liczby analizowanych pokoleń prawdopodobnie spowoduje wykrycie wspólnych przodków głębiej w rodowodzie i wzrost spokrewnienia rodziców, a co za tym idzie wzrost inbredu potomka. Należy pamiętać, że oszacowany inbred jest swoisty dla danej populacji. Co to znaczy? Poziom inbredu oszacowany dla danego osobnika w Polsce może być inny niż poziom inbredu tego samego osobnika np. w Niemczech. Dlaczego? Ponieważ wszystko zależy od tego, jak kompletny i głęboki rodowód został wykorzystany w obydwu oszacowaniach oraz jaką metodą wykonano obliczenia. Każdy kraj ma swoje założenia dotyczące metody szacowania inbredu oraz bazuje na informacji z własnych ksiąg hodowlanych, dlatego obie wartości inbredu są nieporównywalne między sobą. Poziom homozygotyczności danego osobnika powinien być interpretowany w odniesieniu do średniego poziomu inbredu rodzących się zwierząt w populacji, z której pochodzi, aby móc stwierdzić, czy jest on niski, czy wysoki.
Znając już mechanizm inbredu, przejdźmy do jego skutków. Następstwem inbredu jest zwiększenie liczby homozygotycznych miejsc w genomie osobnika, co powoduje obniżenie zmienności produkowanych komórek jajowych i plemników, a w konsekwencji mniejszą różnorodność potomstwa. Wpływa to na obniżenie zmienności genetycznej całej populacji. Kolejnym skutkiem inbredu jest ujawnianie się wadliwych, zwykle recesywnych alleli oraz niekorzystnych genotypów. Nie jest tak, że inbred wpływa na wytworzenie ich na nowo w populacji, one już występują w genomie zwierząt. Jednak zmniejszenie możliwych do utworzenia kombinacji, wynikające z obniżonej zmienności genetycznej komórek rozrodczych będących efektem przekazywanych identycznych alleli od ojca i matki, powoduje, że częściej są one obserwowane w populacji w postaci różnego rodzaju wad i chorób oraz obniżonej żywotności zwierzęcia. Tym najbardziej zauważalnym dla hodowcy skutkiem inbredu jest obniżenie produkcyjności, rozrodu i zdrowotności zwierzęcia w porównaniu ze zwierzęciem, które pochodzi z kojarzenia po niespokrewnionych rodzicach. Zjawisko to nazywamy depresją inbredową. Poronienia, upadki, gorsza płodność czy produkcja to straty ekonomiczne, które ponosi hodowca w związku z inbredem.
W ostatnich latach zaczęto rozdzielać inbred na tzw. inbred stary (ang. ancient inbreeding) i inbred nowy (ang. recent inbreeding). Inbred stary jest definiowany jako poziom homozygotyczności danego osobnika w wyniku kojarzenia krewniaczego jego rodziców, których przodek/przodkowie znajdują się w odległych pokoleniach. Aby w ogóle móc sklasyfikować inbred jako stary, należy dysponować głęboką i kompletną informacją rodowodową. Inbred potomka określany jako stary jest zwykle niższy i związany ze wspólnym przodkiem rodziców znajdującym się w odległym pokoleniu. Dla każdej populacji ta głębokość może być inna, zwykle wynosi około pięciu lub więcej pokoleń. Nowy inbred jest definiowany jako połowa spokrewnienia rodziców, których wspólny przodek występuje płytko w rodowodzie. Co to oznacza? Potomek takiej pary pochodzi z kojarzenia krewniaczego rodziców, którzy są ze sobą blisko spokrewnieni, a wspólny przodek znajduje się nie dalej niż trzy pokolenia wstecz. Poziom homozygotyczności takiego potomka jest zwykle bardzo wysoki, przekraczający nawet 20%. Inbred ma swoje zarówno negatywne, jak i pozytywne strony. Kojarzenie podobnych zwierząt pozwoliło na ujednolicenie populacji pod względem wielu cech, co spowodowało ogromny postęp hodowlany u bydła. Dzięki praktyce chowu wsobnego możemy tworzyć nowe rasy. Jednak intensywny wzrost inbredu, wąska pula męskich reproduktorów, szczególnie w dobie ery genomowej, wpływają na ograniczenie zmienności populacji i przyspieszają przyrost inbredu z pokolenia na pokolenie. Najnowsze badania wskazują na różny wpływ starego i nowego inbredu na depresję inbredową. Nowy inbred ma zdecydowanie większy i bardziej negatywny wpływ na selekcjonowane cechy. Szczególnie obserwuje się ten efekt na wydajność cech produkcyjnych, tj. mleka, białka i tłuszczu, podczas gdy stary inbred nie ma negatywnego wpływu na te cechy. Obserwuje się nawet minimalny pozytywny efekt starego inbredu na te cechy, co może być związane z konsolidacją populacji. Wyniki te zostały potwierdzone w badaniach z wykorzystaniem informacji zarówno rodowodowej, jak i genomowej. Jednak – co podkreślają autorzy tych badań – ważne są pełne i głębokie rodowody. Jedną z zalet informacji genomowej jest możliwość oszacowania rzeczywistego poziomu inbredu oraz uchwycenie wszystkich rekombinacji i segregacji, która nie jest możliwa dla klasycznego inbredu będącego jedynie przybliżeniem. Jedną z hipotez, która zdaje się wyjaśniać fenomen mocniejszego i negatywnego wpływu nowego inbredu, opiera się na oczekiwanym spadku częstości występowania szkodliwych alleli w czasie, co jest wynikiem selekcji. Ponieważ większość tych alleli jest szkodliwa, chów wsobny zwiększa skuteczność selekcji przeciwko tym allelom poprzez zwiększenie homozygotyczności i wypieranie takich alleli, co się nazywa oczyszczaniem populacji. Jednak oczyszczanie populacji jest bardziej prawdopodobne, gdy istnieje silna presja selekcyjna.
Co w takim razie może zrobić hodowca? Przede wszystkim zwracać uwagę na kompletność i głębokość danych rodowodowych zarówno swoich zwierząt, jak i zwierząt, których materiał genetyczny wykorzystuje w stadzie. Podczas zestawiania par do kojarzeń zwracać uwagę na spokrewnienie pomiędzy wytypowaną parą rodzicielską. Im wyższe spokrewnienie rodziców, tym wyższy będzie inbred potomka. Nie dopuszczać do kojarzenia w bliskim pokrewieństwie przynajmniej do czwartego pokolenia wstecz, ponieważ może to wpłynąć na mocniejszy efekt depresji inbredowej obserwowanej u potomka. Monitorować inbred w swoim stadzie, czy nie przyrasta on zbyt szybko z pokolenia na pokolenie. Inbredu urodzonych zwierząt nie da się już cofnąć ani zmienić, można zarządzać inbredem zwierząt, które dopiero się urodzą, poprzez odpowiednie dobory. Aby uzyskiwać dobre wyniki hodowlane i produkcyjne w stadzie, hodowca powinien kierować się zasadą minimalizacji inbredu przy jednoczesnej maksymalizacji celu hodowlanego, a długofalowa perspektywa powinna być częściej wybierana niż krótkofalowe zyski.