
Coraz mniej nosicieli defektów genetycznych wśród buhajów hodowanych w Polsce
Mija 30 lat, odkąd w Polsce rozpoczęto systematyczne badania przesiewowe, pozwalające na wykrywanie buhajów będących nosicielami defektów genetycznych. Badania te pozwoliły na niemalże całkowitą eliminację głównych defektów, ale pojawiają się nowe defekty, których możemy uniknąć poprzez stosowanie oceny genomowej.
tekst: prof. dr hab. inż. Stanisław Kamiński, Uniwersytet Warmińsko-Mazurski w Olsztynie
Katedra Genetyki Zwierząt
Hodowla w czystości rasy, z jaką mamy do czynienia w Polsce w obrębie rasy holsztyńsko-fryzyjskiej, przynosi niekwestionowane korzyści. Jest to przede wszystkim wzrost wydajności produkcji mleka, który poprzez odpowiednią kontrolę użytkowości, selekcję i dobór krów i buhajów tej samej rasy powoduje kumulację pożądanych wariantów genów w kolejnych generacjach. Jeśli konkretny wariant jest korzystny, jego efekt się podwaja, kiedy występuje w postaci homozygotycznej, to znaczy, że został odziedziczony od obojga rodziców. Istnieją jednak geny, których warianty mogą mieć efekt negatywny dla zdrowia i rozwoju zwierzęcia. Te z kolei, kiedy wystąpią w postaci homozygotycznej, mogą pogarszać różne cechy, a niektóre nawet mogą objawić się w sposób bardzo ostry – poprzez wywołanie wad anatomicznych, zaburzeń biochemicznych, dysfunkcji fizjologicznych, a nawet śmierci. Efekty takich genów nazywa się ogólnie wadami lub defektami genetycznymi. Im większy odsetek genów w postaci homozygotycznej (zwany inbredem), tym większe prawdopodobieństwo ujawnienia się defektów genetycznych. Dlatego właśnie hodowle o największej tradycji i stojące na najwyższym poziomie pierwsze rozpoczęły programy identyfikacji, monitorowania i ewentualnej eradykacji (usuwania) nosicieli defektów genetycznych.
Postęp w wykrywaniu nosicieli defektów genetycznych u bydła był możliwy dzięki nowym metodom analizy DNA. Na przełomie wieków XX i XXI stało się możliwe określenie wariantów pojedynczych genów zwierzęcia na podstawie jego DNA, bez oczekiwania na ujawnienie się fenotypu, czyli obserwowanej cechy lub choroby u samego badanego zwierzęcia albo jego potomstwa czy też krewnych. Stało się to w bardzo ważnym okresie dla rozwoju polskiej hodowli bydła mlecznego, czyli w trakcie tzw. holsztynizacji bydła, polegającej na intensywnym krzyżowaniu bydła rodzimej rasy polskiej nizinnej czarno-białej z rasą holsztyńsko-fryzyjską. W tym newralgicznym okresie w olsztyńskiej uczelni utworzono laboratorium biologii molekularnej, pierwsze w Polsce w obszarze nauk o zwierzętach, które podjęło się zadania wykrywania nosicieli mutacji warunkujących defekty genetyczne u bydła mlecznego.
W terminologii genetycznej nosiciel jest osobnikiem zdrowym, nie widać, żeby miał jakieś istotne kliniczne zaburzenia anatomiczne czy fizjologiczne w porównaniu do swoich rówieśników. Tymczasem w ściśle określonym miejscu na konkretnym chromosomie posiada mutację, której szkodliwy efekt ujawnia się tylko wtedy, kiedy od obojga rodziców tę mutację odziedziczy. Jeśli zatem – a tak dzieje się najczęściej – krowa posiadająca normalny wariant genu zostanie skojarzona z buhajem nosicielem, wówczas można się spodziewać, że połowa ich potomstwa będzie nowymi nosicielami, które także nie będą wyróżniać się od swoich rówieśników. Mutacja ta jednak będzie się przenosić w kolejnych generacjach, do momentu aż osobniki dalece spokrewnione, a będące nosicielami, zostaną skojarzone w pary. Tak właśnie dzieje się w hodowli bydła, gdzie wybitny buhaj, poprzez inseminację krów jego nasieniem w wielu krajach, może być przodkiem setek tysięcy potomstwa rozproszonego na całym świecie. Po wielu latach jego potomek będący nosicielem, ze względu na wysoką międzynarodowo uznaną wartość hodowlaną, jest kojarzony z krowami nosicielkami tej samej mutacji (także zdrowymi). W efekcie u 25% potomstwa dochodzi do spotkania się dwóch kopii tej samej mutacji – a to już prowadzi do pełnego ujawnienia się defektu. Zazwyczaj jest to obumieranie zarodków lub płodów albo urodzenia martwych cieląt lub cieląt osłabionych i chorych, które są usuwane przez hodowcę ze stada. Wykrywanie nosicieli defektów daje zatem podwójną korzyść: nie dopuszcza do ujawnienia się szkodliwego efektu mutacji, ale także – co równie ważne – hamuje jej rozprzestrzenianie się.
Pierwszym defektem identyfikowanym w Polsce na podstawie DNA była wczesna obumieralność zarodkowa, znana pod międzynarodowym skrótem DUMPS (Deficiency of UridinomonoPhosphate Synthase – niedobór syntazy urydyno-mono-fosforanowej). Okazało się, że po opracowaniu testu w 1993 roku w ciągu 9 lat przebadano 2,2 tys. buhajów dopuszczonych do rozpłodu i – w przeciwieństwie do sytuacji w innych krajach – w Polsce nie stwierdzono ani jednego nosiciela tego defektu.
Na kolejne defekty nie trzeba było jednak długo czekać. Nasilający się import nasienia czołowych buhajów sprawił, że do polskiej hodowli dotarły także nowe warianty genów wywołujących defekty.
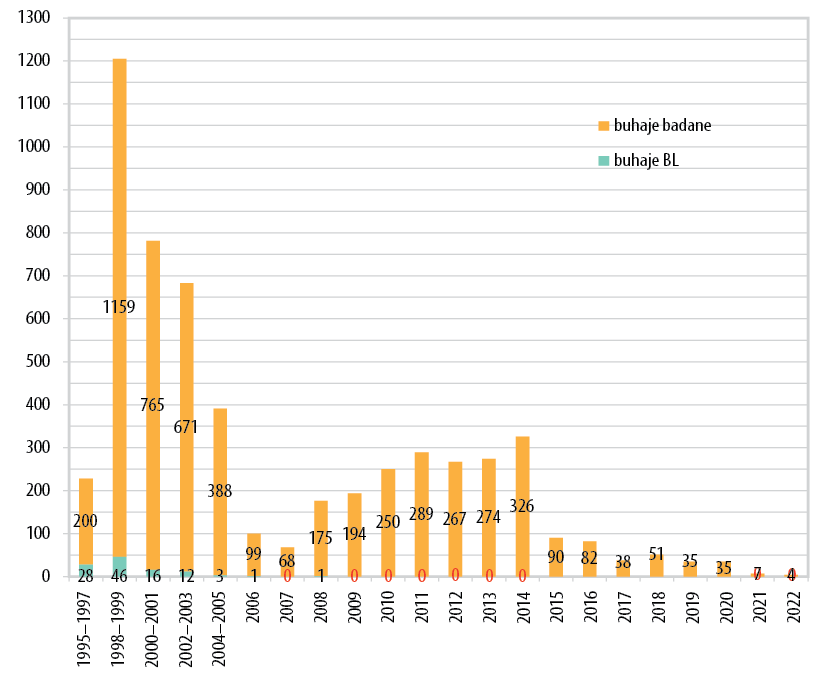
Po badaniach przesiewowych na DUMPS ośrodek olsztyński identyfikował przez wiele lat nosicieli defektu BLAD (BLAD – Bovine Leukocyte Ahesion Deficiency – niedobór adhezji leukocytów bydła), który wywoływał obniżenie odporności cieląt prowadzące do nawracających infekcji układu oddechowego i pokarmowego, a w konsekwencji do gorszego wzrostu i rozwoju, prowadzącego zazwyczaj do wybrakowania lub upadków. Mutacja przyczynowa defektu BLAD nie wywołuje u nosicieli żadnych nieprawidłowości w funkcjonowaniu układu immunologicznego. Opracowanie w UWM w Olsztynie oryginalnej i ekonomicznej metody identyfikacji mutacji wywołującej BLAD otworzyło możliwość badań przesiewowych w Polsce. W celu zapobiegania stratom zwierząt w hodowli masowej od 1999 roku obligatoryjnym testom na nosicielstwo BLAD były poddawane buhajki odmian czarno-białej i czerwono-białej przeznaczone do rozrodu. Skala nosicielstwa BLAD osiągnęła w Polsce w 2000 roku poziom 3,2% wśród buhajów dopuszczonych do rozpłodu, a w próbie populacji masowej krów – aż 4,6%. Strategia systematycznego monitorowania i eradykacji nosicieli BLAD wśród wszystkich młodych buhajów zakupowanych do stacji unasieniania spowodowała, że już od 2009 roku do dziś nie odnotowano ani jednego nowego nosiciela BLAD (rycina 1).
Kiedy liczba nosicieli defektu BLAD malała do jednego na rok, pojawił się nowy defekt CVM (Complex Vertebral Malformation – złożone zniekształcenia kręgosłupa). CVM jest przyczyną zamierania zarodków w różnych okresach ciąży, poronień i urodzeń martwych cieląt. Jak dotąd wykryto tę wadę wyłącznie u bydła rasy holsztyńsko-fryzyjskiej odmiany czarno-białej. Znowu u nosicieli nie występują widoczne oznaki jej działania. Należy oczekiwać natomiast obniżenia płodności krów nosicielek, będącego wynikiem krzyżowania z buhajami nosicielami CVM.
Kojarząc ze sobą heterozygoty, można się spodziewać ok. 25% cieląt obarczonych tą wadą letalną. Badacze duńscy wykazali, że 77% zarodków – homozygot recesywnych z CVM – ulega poronieniu przed 260. dniem ciąży (z tego aż 29% krów roni przed setnym dniem ciąży). Ci sami uczeni zaobserwowali również, że okres międzywycieleniowy u krów zacielonych i noszących zarodki obciążone wadą wydłuża się średnio o 33 dni. Prowadzi to do istotnego spadku wydajności mlecznej, zmniejszenia liczby cieląt i w praktyce może być powodem wcześniejszego brakowania krów.
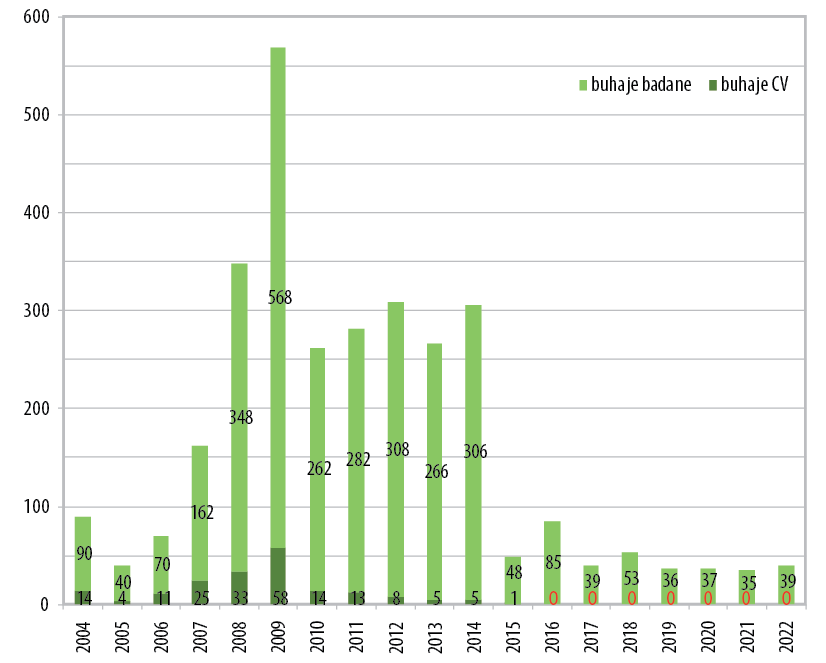
Także w przypadku defektu BLAD nasz ośrodek prowadził systematyczne badania od 2004 roku. Na rycinie 2 widać, że w populacji ponad 1,8 tys. buhajów badanych w latach 2004–2022 wykryliśmy obecność 303 buhajów nosicieli CVM (16,6%). Konsekwentna realizacja programu zalecającego badanie na nosicielstwo wszystkich buhajków sprzedawanych do czterech polskich stacji unasieniania zwierząt spowodowała drastyczny spadek liczby nosicieli od 2010 roku do tego stopnia, że od 2016 nie zaobserwowano w Polsce żadnego nosiciela (rycina 2).
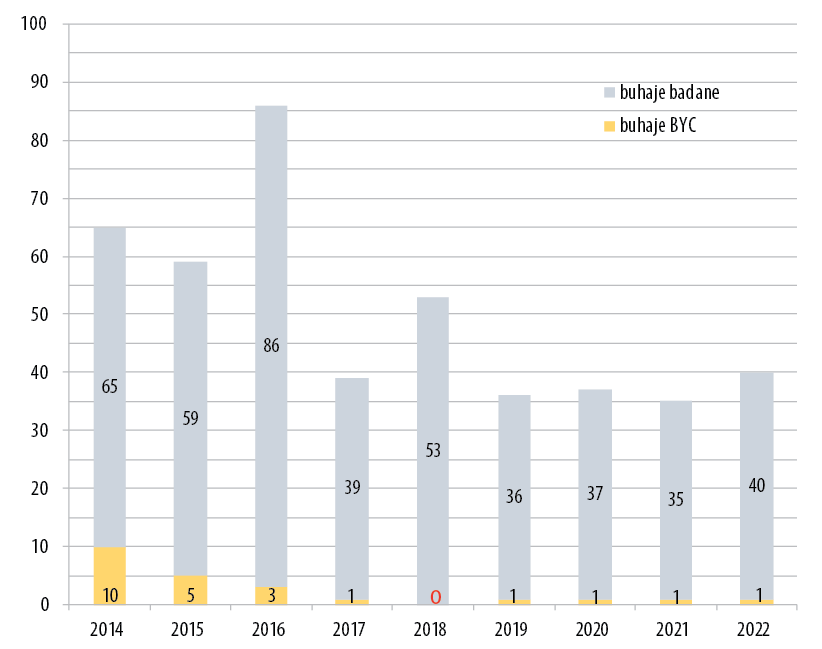
Znowu jednak na kolejny defekt nie trzeba było długo czekać. Mowa o defekcie zwanym Brachyspina, który powodował podobne objawy kliniczne jak CVM. Termin Brachyspina (BY) oznacza skrócony kręgosłup i rzeczywiście jest to główna anatomiczna cecha zwierząt obciążonych tym defektem. Inne symptomy występujące u osobników chorych to poronienia i martwe urodzenia. Skala oznaczeń nosicielstwa była w tym przypadku mniejsza, ale efekt końcowy podobny – po kilku latach identyfikacji nosicieli i usuwania ich z rozpłodu przez ich właścicieli obecnie są wykrywani pojedynczy nosiciele (rycina 3).
Ostatnio do grupy starych defektów dołączyły nowe, tzw. Holsztyńskie Haplotypy (HH). Wykryto je dzięki analizie dziedziczenia markerów SNP używanych w selekcji genomowej. Zauważono bowiem, że niektóre markery występują jedynie w postaci dwóch genotypów u żywych zwierząt (np. AA i AB). Trzeci genotyp (BB) nie pojawiał się u cieląt. Stąd wniosek, że ten genotyp homozygotyczny musi być letalny (śmiertelny). Takie haplotypy mogą przynosić bardzo wymierne straty ekonomiczne. Jeden z ostatnich nowych defektów genetycznych, nazwany HH1, w ciągu ostatnich 35 lat wywołał w USA obumarcie ok. 525 tys. zarodków i spowodował straty na sumę 420 mln dolarów. Wszystko za sprawą buhaja o imieniu Chief, urodzonego w 1962 roku. Był to drugi w całej historii hodowli bydła fryzyjskiego buhaj, który wywarł największy wpływ na całą rasę. Nikt nie wiedział, aż do niedawna, że był nosicielem mutacji wywołującej wczesne zamieranie zarodków.
Obecnie zarówno stare, jak i nowe defekty (z wyjątkiem Brachyspina) są identyfikowane na mikromacierzy i oznaczane skrótami HH (Holstein Haplotype). Przy tej okazji uporządkowano międzynarodowe symbole wszystkich defektów (choć stare symbole nadal są używane w dokumentacji hodowlanej i w katalogach buhajów). Dla defektów wykrytych jeszcze przed upowszechnieniem się oceny genomowej wprowadzono symbole literowe, np. HHB (dla defektu BLAD), HHC (dla defektu CVM), HHD (dla defektu DUMPS) czy HHM (dla syndaktylii). Z kolei dla haplotypu związanego z defektem Brachyspina zarezerwowano symbol HH0. Pozostałych haplotypów mamy aż siedem, oznaczanych od HH1 do HH7.
W systemie Symlek obowiązuje następujący sposób zapisu wyniku testu na HH, np. dla HH7 są trzy możliwe zapisy genotypu: HH7F – (Free, wolny), HH7C – (Carrier, nosiciel), HH7S – (Sick, chory).
Efekty fenotypowe i ekonomiczne, jakie wywołują te haplotypy w stanie homozygotycznym, są zróżnicowane i zależą głównie od ich frekwencji oraz fazy ciąży, w której ujawnia się ich działanie. Im wyższa częstość występowania haplotypu i im późniejsza faza ciąży, tym starty wywołane haplotypami są większe. Do poronień dochodzi zazwyczaj przed 60.–100. dniem ciąży (HH2–HH7) lub we wszystkich stadiach ciąży (HH1).
Trzeba zaznaczyć, że w niektórych krajach negatywny wpływ haplotypów na cechy płodności jest uwzględniany w genomowej ocenie buhajów. Chodzi o dwie cechy – SCR (sire conception rate) i DPR (daughter pregnancy rate). A zatem jeśli buhaj jest homozygotyczny pod względem jednego lub kilku haplotypów, wartość hodowlana dla tych cech powinna być znacząco niższa, co stanowi skuteczny sygnał ostrzegawczy dla hodowcy. W krajach, w których nie ocenia się podobnych cech, pozostaje zwracanie uwagi na oznaczenia buhajów, czy są nosicielami tych haplotypów. Informację tę znajdziemy w dokumentacji rodowodowej lub marketingowej (np. w katalogach buhajów).
Wszystkie HH (z wyjątkiem Brachyspina) znajdują się na mikromacierzy Illumina, stosowanej do oceny genomowej bydła na świecie, także w Polsce (w ramach Spółdzielni EuroGenomics). Decydując się na zgenotypowanie jałówki lub buhajka, pozyskujemy nie tylko ich szacowaną wartość genomową dla cech użytkowych, ale także informację, czy są nosicielami defektów genetycznych.
Problemu występowania defektów genetycznych nie można ani trywializować, ani nadmiernie eksponować. Nosiciele często były wybitnymi rozpłodnikami i dlatego dopuszczane były do rozrodu przez firmy inseminacyjne. Szkody wynikające z poronień lub martwo urodzonych cieląt w skali populacji mogły być mniejsze niż korzyści, jakie przynosiło dla postępu genetycznego, np. w zakresie produkcji mleka, użycie takiego buhaja. Z kolei całkowite lekceważenie statusu nosicielstwa u buhaja jest niewskazane. Fakt wykrycia nosicielstwa powinien wywołać obowiązek monitorowania potomstwa nosiciela, zwłaszcza męskiego kwalifikowanego do rozpłodu, ale także matek buhajów i dawczyń zarodków. W praktyce, przy ogromnej konkurencji firm inseminacyjnych, buhaje nosiciele defektów szkodzą wizerunkowi firmy i dlatego są z reguły usuwane z oferty nasienia, bez zbędnego rozgłosu.
W niektórych krajach (np. Francji czy Danii) funkcjonują programy wczesnego wykrywania nowych defektów i eliminowania osobników będących nosicielami. Czołowe hodowle światowe po wykryciu pierwszych zdeformowanych cieląt, wykazujących anomalie anatomiczne, rejestrują je, opisują i sprawdzają, czy cielęta te mają wspólne pochodzenie. Jeśli stwierdzą, że mają wspólnego przodka, podejmują badania prowadzące do identyfikacji tzw. mutacji sprawczej, a następnie opracowują test genetyczny na wykrywanie nosicieli. Zazwyczaj taki test podlega opatentowaniu. W tym czasie oczyszczają swoją populację z nosicieli tego właśnie defektu. Zagraniczni hodowcy w tym samym czasie nieświadomie kontynuują inseminację nasieniem buhajów nosicieli. W efekcie nosiciele przenikają do krajów importujących, a Polska do takich cały czas należy. Byłoby wskazane, aby Polska wdrożyła własny program wczesnego wykrywania cieląt dotkniętych defektami. Hodowca, który zgłasza przypadki defektów, polepsza jakość całej populacji. Do takich standardów musimy dążyć.








