Jak pokryć zapotrzebowanie krów mlecznych na witaminy? Część 2.
Witaminy rozpuszczalne w tłuszczach, w tym m.in. A, D i E, mają bardzo duży wpływ na prawidłowe funkcjonowanie organizmu krów i ich potomstwa. Dlatego warto zadbać o ich odpowiedni poziom w dawkach pokarmowych.
TEKST: PROF. ZYGMUNT M. KOWALSKI, UNIWERSYTET ROLNICZY W KRAKOWIE; ZDJĘCIA: RYSZARD LESIAKOWSKI
W części 1., w poprzednim numerze „Hodowli i Chowu Bydła”, omówiłem podstawowe zasady żywienia witaminowego krów mlecznych. Chociaż mikroorganizmy żwacza (bakterie i pierwotniaki) produkują znaczne ilości witamin z grupy B, w tym biotynę i witaminę K, to w niektórych sytuacjach ta ilość może być niewystarczająca. Na przykład w sytuacji wysokich wydajności mleka i związanego z tym wysokiego zapotrzebowania na witaminy, ilość witamin produkowanych w żwaczu może być za mała. Ponadto w żwaczu krów żywionych znacznymi ilościami pasz treściwych – co wymusza wysoka wydajność mleka – utrudnione jest namnażanie się bakterii i pierwotniaków, które odpowiadają za syntezę witamin, zwłaszcza biotyny.
Z kolei pokrycie zapotrzebowania krów na witaminy rozpuszczalne w tłuszczach (A, D3 i E) zależy z jednej strony od ich zawartości w dawkach pokarmowych, a z drugiej strony od pobrania paszy. O ile coraz rzadziej spotyka się hodowców „oszczędzających” na mieszankach mineralno‑witaminowych, o tyle nie wszyscy stosują je właściwie, dobierając odpowiednią mieszankę do stanu fizjologicznego krowy. Ponadto krowa, która nie pobiera wystarczającej ilości suchej masy w okresie przejściowym, nie pobiera również witamin, co może być powodem niepokrywania zapotrzebowania na te ważne składniki pokarmowe.
W części 1. zwróciliśmy uwagę także na fakt, że żywienie witaminowe krów nie polega jedynie na stosowaniu takich dawek witamin, które pozwolą na uniknięcie objawów niedoboru. Przez uzupełnianie dawek pokarmowych witaminami, w ilościach które przekraczają fizjologiczne zapotrzebowanie organizmu krowy, wpływa się na jej produkcyjność i zdrowie. To przekraczanie zapotrzebowania powinno opierać się na zaleceniach wynikających z badań naukowych, a nie wynikać z chęci większej sprzedaży mieszanki mineralno-witaminowej czy dodatku paszowego. W niniejszym artykule omówię rolę najważniejszych witamin rozpuszczalnych w tłuszczach dla krów mlecznych, a także praktyczne aspekty ich stosowania.

Witamina A i β-karoten
Witamina A, zwana retinolem, należy do witamin rozpuszczalnych w tłuszczach. Bierze udział w budowie i regeneracji naskórka i nabłonka, jest istotna dla wzrostu, płodności oraz odporności, bo stymuluje wytwarzanie przeciwciał. Jej brak powoduje zmiany na skórze i błonach śluzowych, zahamowanie wzrostu, zaburzenia płodności i zwiększoną podatność na choroby. Witamina A nie występuje w paszach roślinnych, co wymusza konieczność jej uzupełniania w dawkach dla krów. Chociaż retinol łatwo ulega utlenianiu, np. w czasie jełczenia tłuszczów, witamina syntetyczna stosowana w mieszankach mineralno-witaminowych jest otoczkowana, co zmniejsza ryzyko utleniania. W paszach roślinnych występuje prowitamina witaminy A, czyli β-karoten, także dobrze rozpuszczalny w tłuszczach, a nierozpuszczalny w wodzie. Podobnie jak witamina A, β-karoten równie łatwo się utlenia.
Bogatym źródłem β-karotenu są pasze zielone, zwłaszcza młode, bogato ulistnione, a także marchew. Konserwacja zielonek, np. produkcja siana na pokosach, przy złej pogodzie powoduje straty prawie całego β-karotenu, głównie z powodu działania promieni słonecznych, wymywania przez deszcz, a także obłamywania liści. Ta ilość β-karotenu, która uniknęła strat w czasie suszenia, zostanie utracona w czasie przechowywania siana. W trakcie produkcji kiszonek z traw czy roślin motylkowatych następują również straty β-karotenu, sięgające nawet do 80%, zwłaszcza przy długim okresie przewiędnięcia lub podsuszania materiału przed kiszeniem. Ziarna zbóż, z wyjątkiem kukurydzy i pszenicy oraz ich pochodnych, są praktycznie pozbawione β-karotenu.
Krowy pobierają witaminę A w mieszankach mineralno-witaminowych lub w premiksach, a także uzyskują ją z przemian β-karotenu. Znaczna część witaminy A jest rozkładana przez bakterie w żwaczu (około 70%), natomiast β-karoten jest bardziej odporny na procesy rozkładu bakteryjnego, bo bakterie „niszczą” około 25% tej prowitaminy. Przy niskim pH płynu żwacza większa część β-karotenu (nawet 50–80%) podlega procesom rozkładu bakteryjnego. Krowy z nadmierną kwaśnością płynu żwacza (SARA) narażone są więc na wieksze ryzyko niedoborów witaminy A i β-karotenu.
W błonie śluzowej jelita cienkiego z 1 cząsteczki β-karotenu powstają 2 cząsteczki witaminy A. Z 1 mg β-karotenu powstaje około 400 j.m. witaminy A. Pozostała część β-karotenu, która nie przekształciła się w witaminę A, zostaje wchłonięta do limfy, z której przedostaje się do krwi, a wraz z nią do wątroby oraz do gruczołu mlekowego i następnie do mleka. Wchłanianie β-karotenu jest dużo gorsze niż wchłanianie witaminy A.
Witamina A po wchłonięciu w jelitach transportowana jest do wątroby, gdzie może być magazynowana. Także wchłonięty β-karoten jest magazynowany w wątrobie i służy do syntezy witaminy A. Zapasy witaminy A w wątrobie krowy wykorzystywane są na przykład w okresie produkcji siary, która zawiera około 10–100 razy więcej witaminy A niż mleko. W tym kontekście tak ważne jest podawanie krowom zasuszonym odpowiednich ilości witaminy A. Szczególnie „cenne” są zapasy witaminy A i β-karotenu zgromadzone w wątrobie w okresie żywienia zielonkami. Mogą być wykorzystywane w czasie 4–8 miesięcy.
Jedną z podstawowych funkcji witaminy A jest udział w procesach wzrostu komórek nabłonka. Jest niezbędna dla utrzymania prawidłowej struktury i czynności komórek nabłonkowych skóry i błon śluzowych. Niedobory witaminy A powodują degenerację komórek nabłonkowych organów układu oddechowego, pokarmowego, moczowego i rozrodczego (np. rogowacenie skóry błony śluzowej pochwy). Suchy, skeratynizowany nabłonek nie stanowi bariery dla infekcji bakteryjnych, wirusowych i pasożytniczych. Niedobory witaminy A sprzyjają więc infekcjom układu pokarmowego, oddechowego oraz rozrodczego.
Rola witaminy A w rozrodzie krów nie jest jednoznaczna, ale często uważa się ją za niezbędną dla dobrego rozrodu krów. Witamina A wpływa na syntezę hormonów steroidowych, a przez to na funkcje rozrodcze. Brak tej witaminy może powodować zaburzenia w rozrodzie, w tym zmniejszenie wskaźnika zapładnialności. Szczególnie groźne mogą być niedobory w okresie późnej ciąży, mogące powodować poronienia, zatrzymywanie łożyska, oraz zmniejszające masę urodzeniową cieląt. Cielęta rodzą się słabsze, często ślepe i martwe. W przypadku niedoborów w początku ciąży zapłodnione komórki jajowe trudniej implantują się w błonie śluzowej macicy. Źle odżywiony embrion zamiera lub ulega deformacji. Niedobory witaminy A we wczesnej ciąży mogą więc być powodem utraty ciąży.
Znacznie częściej pisze się o pozytywnym wpływie β-karotenu na wskaźniki rozrodu, chociaż efekty uzupełniania dawek zależą bardzo mocno od poziomu β-karotenu we krwi u zwierząt niedokarmianych. Efekt dokarmiania jest widoczny zwłaszcza wtedy, gdy w grupie kontrolnej stężenie β-karotenu we krwi jest poniżej 200–300 µg/100 ml.
Znaczne ilości β-karotenu znajdują się w ciałku żółtym krowy. Może to być powodem poprawy płodności u krów otrzymujących dodatek β-karotenu do dawek zawierających wystarczające ilości witaminy A. U krów dokarmianych dodatkiem β-karotenu obserwowano jego pozytywny wpływ na zwiększenie wskaźnika zapładnialności, skrócenie okresu międzyciążowego, poprawę wskaźnika skuteczności pierwszego unasiennienia oraz na zmniejszenie częstotliwości występowania cichych rui i cyst jajnikowych. Zmniejszała się również częstotliwość wczesnych poronień. Wskazuje to na dodatkową rolę β-karotenu w rozrodzie krów, poza jego rolą jako prowitaminy witaminy A. Z tego powodu nie traktuje się obecnie β-karotenu jedynie jako prekursora witaminy A.
Hodowców interesuje także znaczenie witaminy A oraz β-karotenu w profilaktyce zapalenia gruczołu mlekowego. Wyniki badań z tego zakresu nie są jednak jednoznaczne. Podobnie jak w przypadku zaburzeń w rozrodzie, reakcja krów na dodatek witaminy A czy β-karotenu zależy od stopnia pokrycia zapotrzebowania bez uzupełniania.

Ile witaminy A i β-karotenu w dawce pokarmowej?
W większości systemów żywienia krów zapotrzebowanie na witaminę A odnosi się do pobrania suchej masy (SM). Najczęściej proponuje się, w zależności od okresu laktacji, zapotrzebowanie od 3200 do 4000 j.m./kg
pobranej SM. Tak więc krowa pobierająca 25 kg SM powinna pobrać od 80 000 do 100 000 j.m. witaminy A dziennie, co odpowiada 200–250 mg
β-karotenu/dzień. W 1 kg mieszanki mineralno-witaminowej powinno być około 1 000 000 j.m. witaminy A,
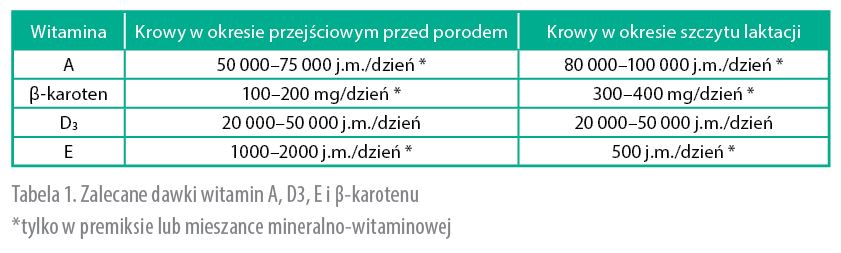
gdy dawka mieszanki wynosi 100 g/dzień. Gdy jest wyższa, zawartość witaminy A powinna być niższa (np. 500 000 j.m., gdy dawka mieszanki wynosi 200 g/dzień). Dotychczas w ustalaniu zapotrzebowania na witaminę A nie uwzględnia się wydajności mleka, bo ta zależy od pobrania SM. W okresie zasuszenia krowa powinna pobierać około 50 000 do 75 000 j.m. witaminy A/dzień.
Chociaż wpływ β-karotenu na wskaźniki rozrodu nie jest jednoznaczny, powszechną praktyką stało się uzupełnianie dawek pokarmowych β-karotenem. Najbardziej popularne zalecenia to dodatek 300–400 mg β-karotenu w pierwszych 90 dniach laktacji, zwłaszcza wtedy, gdy poziom β-karotenu we krwi jest niski. W sytuacjach szczególnych (mastitis!) zaleca się nawet podawanie 600 mg β-karotenu dziennie. Są to dawki dużo większe niż zalecenia dotyczące ilości β-karotenu niezbędnego do syntezy witaminy A. Praktyka zwiększania dawek dziennych β-karotenu ma w USA czy w Europie Zachodniej wielu zwolenników. NRC [2001] zaleca uzupełnianie dawek w β-karoten, gdy: krowy żywione są niskiej jakości paszami objętościowymi, w dawce dominuje kiszonka z kukurydzy, znajduje się w niej dużo pasz treściwych, jest zwiększona ilość patogenów w środowisku krowy oraz w sytuacji zwiększonej immunosupresji (okres okołoporodowy).
Ze względu na bardzo zmienną zawartość β-karotenu w paszach w praktycznym żywieniu normy NRC (2001) proponują nieuwzględnianie w dawkach pokarmowych zawartości β-karotenu oraz witaminy A. Całość zapotrzebowania pokrywa witamina A oraz β-karoten dostarczone w mieszankach mineralno-witaminowych lub premiksach. Krowy zasuszone powinny pobierać 100–200 mg β-karotenu/dzień. Uzasadnione jest rozpoczęcie uzupełniania dawek w β-karoten 2–3 tygodnie przed porodem.
Witamina D
Witamina D również należy do witamin rozpuszczalnych w tłuszczach. Jej dwie formy to cholekalcyferol (witamina D3) i ergokalcyferol (witamina D2). Witamina D3 znajduje się w paszach pochodzenia zwierzęcego, natomiast witamina D2 – w paszach roślinnych, w tym na przykład w sianie suszonym na słońcu. Witamina D3 jest odporna na działanie temperatury oraz niskiego i wysokiego pH i jest mniej podatna na utlenianie niż witamina A. W mieszankach mineralno‑witaminowych stosuje się witaminę D3 syntetyzowaną chemicznie.
Pod wpływem działania promieni słonecznych (UV) ergosterol zawarty w paszach roślinnych przekształcany jest w witaminę D2. Im większe jest działanie słońca na zielonkę suszoną na siano, tym więcej zawiera ono witaminy D2. Więcej witaminy D2 znajduje się więc w kiszonkach z materiału przewiędniętego i podsuszonego niż z materiału świeżego. Promieniowanie UV jest także niezbędne dla syntezy witaminy D3 z 7-dehydrocholesterolu znajdującego się w skórze i naskórku zwierząt. Skoro nie przechodzi przez szkło okienne, zwierzęta utrzymywane alkierzowo mają ograniczone możliwości syntezy witaminy D3.
U przeżuwaczy witamina ta jest bardziej biologicznie aktywna niż witamina D2, która w znacznym stopniu rozkładana jest w żwaczu (nawet do 75%), zwłaszcza przy żywieniu dawkami z dużym udziałem pasz objętościowych. Witamina D2 wchłania się w jelitach podobnie łatwo, jak witamina D3. Wątroba bydła magazynuje witaminę D3 i D2, i te zapasy wystarczają u dorosłej krowy na kilka miesięcy.
Witamina D3, po wchłonięciu w dwunastnicy i dostaniu się do wątroby (do niej dostaje się również witamina D3 powstająca w skórze), przekształcana jest w 25-hydroksycholekalcyferol, a ten dalej przekształcany jest w nerkach w aktywną formę witaminy D3 czyli w 1,25-dihydroksycholekalcyferol. Ta forma transportowana jest przez krew do tkanek, w tym do jelit i kości. Syntezę w nerkach kontrolują parathormon, produkowany przez przytarczyce, oraz kalcytonina, hormon tarczycy. Gdy poziom Ca we krwi jest niski, przytarczyce wydzielają więcej parathormonu, co zwiększa produkcję aktywnej witaminy D3, która zwiększa wchłanianie Ca w jelitach i hamuje jego wydalanie z moczem.
Witamina D3 bierze udział w regulacji poziomu Ca i P w organizmie, a zwłaszcza w regulacji ich wchłaniania z przewodu pokarmowego, wydalania przez nerki i odkładania w kościach. Objawy niedoboru witaminy D3 wiążą się z zaburzeniami gospodarki Ca i P, co w przypadku krów mlecznych dotyczy przede wszystkim hipokalcemii w okresie porodowym.
Gdy koncentracja Ca we krwi jest niska, zwiększa się koncentracja aktywnych form witaminy D3 w błonie śluzowej jelita, a przez to wzrasta wchłanianie Ca. W sytuacjach niedoboru Ca zwiększa się jego wtórna resorpcja w nerkach i zmniejsza wydalanie z moczem. Ważna jest rola witaminy D3 w prewencji zalegania poporodowego u krów mlecznych, bo ułatwia uruchamianie Ca z kości. Witamina D3 odpowiada także za prawidłowe funkcjonowanie systemu odpornościowego krowy.
W normalnych warunkach żywieniowych rzadko obserwuje się objawy niedoboru witaminy D u krów mlecznych. Zwierzęta te dodatkowo magazynują znaczne ilości witaminy D w wątrobie, gdy jej pobranie w paszy przewyższa zapotrzebowanie. Zapasy te mogą dostarczać witaminy D w okresach niedoborowych, trwających nawet kilka tygodni. Okresowe niedobory witaminy D mogą występować przy alkierzowym utrzymywaniu krów. Obserwuje się wtedy zaburzenia w cyklu rujowym, zwłaszcza brak rui po porodzie. Duża zmienność w zawartości witaminy D w paszach, a także zmienne warunki utrzymania zwierząt powodują, że zaleca się uzupełnianie dawek dla cieląt, krów mlecznych oraz młodego bydła opasowego dodatkami zawierającymi witaminę D, zwłaszcza że należy ona do najtańszych witamin.
Witamina E
Witamina E należy również do witamin rozpuszczalnych w tłuszczach. Występuje w paszach zielonych, a ponadto w kiełkach zbożowych, nasionach roślin oleistych, głównie w postaci α, β i γ-tokoferoli, z których najbardziej aktywny biologicznie jest α-tokoferol. Spożycie witaminy E w paszach zielonych zwykle przewyższa zapotrzebowanie na tę witaminę, jednakże znaczna jej część rozkładana jest w żwaczu. W żywieniu dawkami z kiszonką z kukurydzy jako paszą podstawową, a także w określonych stanach fizjologicznych (okres przejściowy przed porodem) istnieje konieczność zwiększenia dawki witaminy E, zwłaszcza w terenach niedoborowych w selen (Se).
Witamina E dość łatwo rozkłada się w żwaczu, zwłaszcza przy niskim pH. Także działanie temperatury (zagrzewanie się kiszonek), przedłużone suszenie zielonek na siano (działanie słońca) lub mielenie pasz znacznie zmniejszają zawartość i aktywność witaminy E. Suszenie na siano może powodować nawet do 90% jej strat. Niewielkie straty w zawartości witaminy E powoduje sam proces kiszenia. Z wyjątkiem nasion roślin oleistych, większość pasz treściwych zawiera mało witaminy E. Ziarna zbóż oraz śruty poekstrakcyjne zawierają jej znacznie mniej niż zielonki. Podgrzewanie pasz treściwych i ich granulowanie znacznie zmniejszają jej koncentrację.
Witamina E jest wchłaniana w jelicie cienkim. Pewne jej ilości są magazynowane w wątrobie. Witamina E odgrywa w organizmie krowy funkcję głównego naturalnego antyoksydanta (przeciwutleniacza) w procesach utleniania wewnątrzkomórkowego. Blokuje utlenianie nienasyconych kwasów tłuszczowych, bo blokuje tworzenie nadtlenków, które hamują działanie wielu enzymów i uszkadzają błony komórkowe. Na szkodliwe działanie nadtlenków najbardziej narażone są komórki mięśni szkieletowych, mięśnia sercowego, komórki wątroby oraz krwi (leukocyty). Witamina E stymuluje działanie peroksydazy glutationu, enzymu zawierającego selen i niszczącego nadtlenki. Współdziała więc z selenem w ochronie błon komórkowych przed utlenianiem powodowanym działaniem wolnych rodników (nadtlenki).
Szczególnie istotna jest rola witaminy E w profilaktyce zatrzymania łożyska, uważanego obecnie za schorzenie spowodowane wadliwym działaniem systemu odpornościowego. Dodatki witaminy E oraz selenu stosowane są dla zapobiegania zatrzymywania łożyska, stanów zapalnych macicy oraz powstawania cyst jajnikowych i skrócenia czasu inwolucji macicy, zwłaszcza w terenach niedoborowych w selen (tj. gdy zawartości Se w roślinach pastewnych wynosi poniżej 0,05 ppm (w SM)).
W okresie okołoporodowym obserwuje się bardzo często zbyt niskie stężenie witaminy E we krwi, co ma konsekwencje zarówno dla zdrowia krowy, jak i cielęcia. Warto tutaj wspomnieć, że tylko niewielka ilość witaminy E przechodzi przez łożysko. Nowo urodzone cielę w całości pokrywa zapotrzebowanie przez spożycie siary i zawartość w niej witaminy E decyduje o zdrowotności cieląt.
Szczególną uwagę zwraca się obecnie na rolę witaminy E i selenu w profilaktyce mastitis. Ochraniają one ciała odpornościowe (neutrofile) przed działaniem wolnych rodników, a neutrofile są podstawą systemu odpornościowego gruczołu mlekowego oraz macicy. Niedostateczna ilość witaminy E we krwi (i selenu) może mieć więc związek ze zwiększoną zapadalnością krów na mastitis oraz na stany zapalne błony śluzowej macicy w kilku pierwszych tygodniach po porodzie. Szczególnie istotne jest spożycie witaminy E przez krowy w okresie zasuszenia. Na przykład podawanie jej w dawkach powyżej 15 j.m./kg SM, tj. powyżej 100 j.m./dzień, a często nawet powyżej 1000 j.m./dzień, wraz z dodatkiem selenu (0,1 lub 0,3 mg Se/kg SM), znacznie zmniejszało ryzyko zapalenia wymienia.
Ile witaminy E w dawce pokarmowej?
We współczesnych zaleceniach dotyczących witaminy E uwzględnia się istotną jej rolę dla systemu immunologicznego, a także fakt bardzo dużej zmienności w jej zawartości w paszach. Aktualnie stosowane normy na witaminę E (np. NRC, 2001) odnoszą się jedynie do witaminy E podawanej w premiksie lub mieszance mineralno-witaminowej. Według NRC zapotrzebowanie krów zasuszonych na witaminę E wynosi 80 j.m./kg SM. Krowa zjadająca 15 kg SM powinna otrzymywać 1200 j.m. witaminy E. W okresie przejściowym (ostatnie 3 tygodnie przed porodem) jej dawka może wynosić nawet 2000 j.m./dzień. Takie pobranie pozwala na utrzymanie bezpiecznego poziomu witaminy E we krwi, co sprzyja profilaktyce mastitis oraz zapobiega zaburzeniom w rozrodzie. Szczególnie podwyższone dawki witaminy E powinny otrzymywać krowy w tych stadach, w których pomimo dobrej profilaktyki mastitis liczba komórek somatycznych w mleku jest bardzo duża. Siara krów żywionych na tym poziomie zawiera dużo witaminy E, co jest istotne dla zdrowia cieląt.
Z kolei zapotrzebowanie krów w laktacji na witaminę E wynosi 20 j.m./kg pobranej SM. Krowa zjadająca 25 kg SM powinna otrzymywać 500 j.m. witaminy E/dzień. Jeżeli mieszanka mineralno-witaminowa skarmiana jest w ilości 100 g dziennie, to w 1 kg powinna zawierać 5000 j.m. witaminy E.
Różnice w zapotrzebowanie na witaminy u krów będących w okresie przejściowym przed porodem oraz krów w okresie laktacji powodują konieczność stosowania innych mieszanek mineralno-witaminowych w okresie zasuszenia („silniejsze”) oraz w laktacji („słabsze”). Ciekawe, jak realizują te wymagania hodowcy, którzy nie grupują krów lub utrzymują krowy przed porodem w sektorach z krowami w laktacji? W tabeli 1. przedstawiono podstawowe zalecenia dotyczące zawartości witamin A, D i E w dawkach dla krów zasuszonych oraz w laktacji.